牛乳中脂肪酶活力检测方法与影响因素研究
2021-08-06 15:53:13 来源: 食品安全导刊
陈柏锡,黄艳艳,曾燕萍,李 斌,邹兰钰,彭远霞,张 伟
(四川新希望乳业有限公司洪雅阳平分公司,四川眉山 620000)
摘 要:原料乳中的某些嗜冷菌会在代谢过程中产生耐热脂肪酶,较高的脂肪酶活力会破坏牛乳中的脂肪球结构并产生多种脂肪酸,游离的脂肪酸易被氧化,产生异味。本文阐述一种利用脂肪酶分解底物产生具有特定吸收波长的物质实现准确检测牛乳中脂肪酶活力的方法,探究了原料奶的运输时间、脱冷储运对脂肪酶活力的影响,以及脂肪酶活力对UHT灭菌乳风味与货架期的影响,期望为相关人员提供一定的参考依据。(四川新希望乳业有限公司洪雅阳平分公司,四川眉山 620000)
关键词:乳脂肪酶活力;比色法检测;原奶储运;UHT灭菌乳
超高温灭菌乳(Ultra High Temperature,UHT乳)经过短时高温的灭菌后,产品达到商业无菌的状态,因此UHT乳具有保质期长、可常温储运等特点[1]。但超高温灭菌乳在储存过程中会出现风味异常、脂肪上浮等问题,严重影响产品的品质和货架期,该现象的主要原因是由UHT乳中脂肪酶导致的。牛乳中的脂肪酶主要有两种来源,一种是天然存在于生乳中,称为天然脂肪酶;另一种是牛乳中的微生物代谢所产生,称为微生物脂肪酶[2]。牛乳中的微生物脂肪酶主要来源于嗜冷菌,虽然嗜冷菌本身的繁殖不会严重影响牛乳的稳定性,且在较为温和的热处理中(如巴氏杀菌)也容易将其杀死,但某些嗜冷菌的代谢产物是耐热脂肪酶,耐热脂肪酶在经过巴氏杀菌甚至超高温灭菌后,仍不能被完全灭活。残留的脂肪酶会分解牛乳中的脂肪,造成脂肪上浮和风味异常,严重破坏灭菌乳的口感和风味,导致灭菌乳的货架期大幅缩短[3]。所以需要一种能够快速、准确且价廉的检测方法检测原料乳中脂肪酶的活力以及探究牛乳中脂肪酶活力的变化规律。
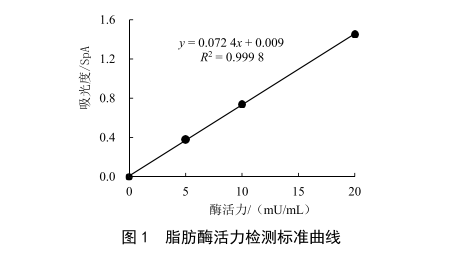
4-硝基苯基辛酸酯(PNP-C)在脂肪酶的作用下会产生在316 nm处有吸光度的物质,根据比尔-朗博定律,在一定浓度下,吸光度大小与吸光度物质浓度呈正比,实验在底物浓度相同的情况下,产生吸光物质浓度与脂肪酶活力成正比。本实验使用脂肪酶标准品配制不同浓度的脂肪酶溶液,分解底物产生吸光度后标定曲线,以此测定样品中脂肪酶活力。
1 材料与方法
1.1 材料与仪器
1.1.1 材料
样品来源于HN、ZX、ZN和XN共4家牧场生乳;
5 U/mL脂肪酶溶液、5 mmol/L4-硝基苯基辛酸酯底物溶液、50 mmol/L pH7.2磷酸缓冲液、终止液:将乙腈与甲酸按照12∶1的比例混匀即可作为终止液。
1.1.2 仪器
紫外分光光度计、水浴加热器、冷冻离心机、移液枪、移液管、水相滤头(0.22 μm)等。
1.2 实验步骤
将5 U/mL脂肪酶溶液配制成100 mU/mL的脂肪酶稀释液备用,分装好的底物在室温下用流水解冻。取20 mL原料奶于50 mL离心管中,于10 000 r/min离心脱脂15 min。离心后将上层脂肪撇去,吸取100 μL清液于900 μL磷酸缓冲液中,混匀,制成1 mL样品处理液。分别吸取300 μL样品处理液于两个2 mL离心管中,其中一个为加标样,并向两个离心管中分别加入180 μL和120 μL磷酸缓冲液,并向加标样的离心管中加入60 μL 100 mU/mL脂肪酶稀释液,最后向离心管中各加入120 μL底物溶液,制成待测样品和加标样品。
制备标准曲线:取4个2 mL离心管,每个离心管加入480 μL、450 μL、420 μL和360 μL磷酸缓冲液,向离心管中加入120 μL底物和0 μL、30 μL、60 μL和120 μL脂肪酶稀释液,混匀后即制成0 mU/mL、5 mU/mL、10 mU/mL和20 mU/mL的标准曲线。将样品和标准曲线置于44 ℃水浴加热1 h取出,向每个离心管中加入1.3 mL终止液,摇匀,即可上机检测。
1.3 标准曲线与数据处理
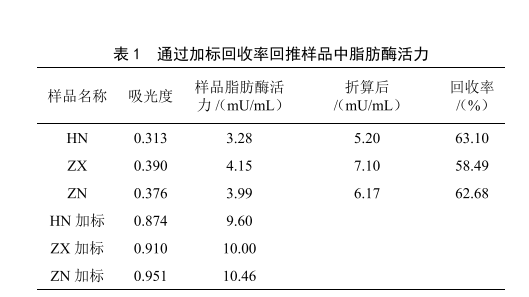
标准曲线见图1,样品中脂肪酶活力见表1。由HN、ZX、ZN 3批原料奶的吸光度计算出样品中的脂肪酶活力,再根据加标回收率折算出样品中脂肪酶活力准确值。需注意的是,样品的回收率应在50%以上(如表1),当回收率偏低时,有可能是离心效果不佳,导致脂肪剩余过多导致的,其原因是样品中的脂肪酶不仅会与底物反应,还会与牛乳中剩余的脂肪反应,在相同的反应时间内,剩余脂肪的含量越高,导致产生吸光物质的浓度比理论值越低,影响检测结果。原料乳离心脱脂率通常为90%~95%。为补偿剩余脂肪产生的影响,每个样品必须进行平行试验和加标回收。
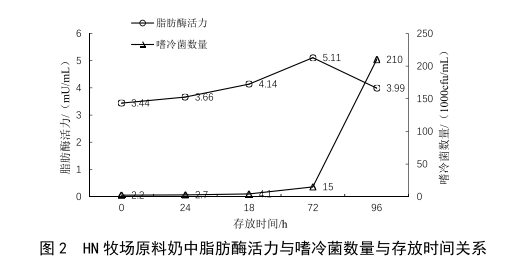
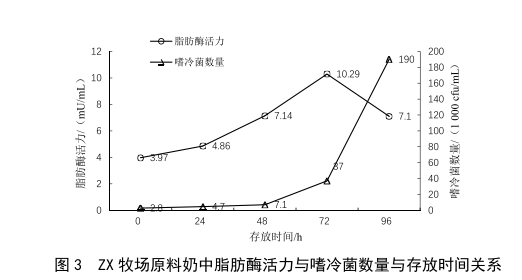
2.1 原奶储运时间对脂肪酶活力的影响
牛乳中耐热脂肪酶是部分嗜冷菌的代谢产物,在冷藏条件下,随着嗜冷菌的繁殖与代谢,牛乳中的脂肪酶活力应随之升高。本次实验取两个不同牧场的原料奶各5份,置于2~6 ℃条件下冷藏避光保存,每隔24 h对其脂肪酶活力和嗜冷菌数量进行检测,绘制出的结果见图2、
图3。
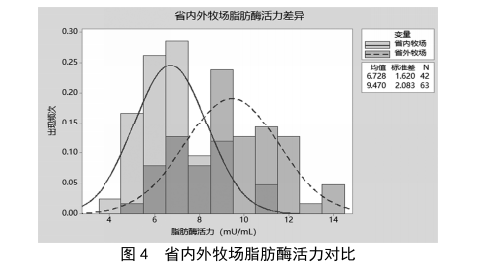
在乳品生产加工的供应链体系中,原料奶的运输时长会随着牧场与工厂距离的增加而增加,跨省牧场的运输时间在20 h以上,运输温度均在4 ℃,原奶运输过程可近似理解为低温储存的过程,本次实验对省外牧场与省内牧场的原料奶进行脂肪酶检测,并统计其数据,结果见图4。由图4可知,省外牧场与省内牧场的原料奶中脂肪酶活力均基本呈现正态分布,省外牧场的脂肪酶活力多数在8~11 mU/mL(平均值9.47 mU/mL),省内牧场的脂肪酶活力普遍在5~8 mU/mL(平均值6.73 mU/mL),省外牧场奶源的脂肪酶活力明显高于省内牧场奶源,与原奶冷藏实验得到的结论一致。
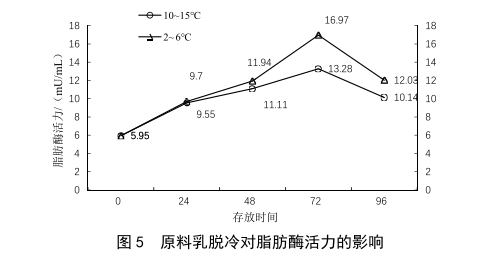
奶车运输和原奶在工厂的奶仓中储存均在4 ℃左右,当奶车运输或原奶储存出现脱冷时(10~15 ℃),也会对原料奶的质量造成影响[4],取相同批次的XN牧场原料乳,分别在2~6 ℃与10~15 ℃环境中保存,模拟奶车正常运输与脱冷的状态,其脂肪酶检测结果如图5所示。当运输或储存出现脱冷时,脂肪酶活力上升的速度也明显增加。在最高点时(72 h),脱冷条件下的脂肪酶活力也明显高于冷藏条件。
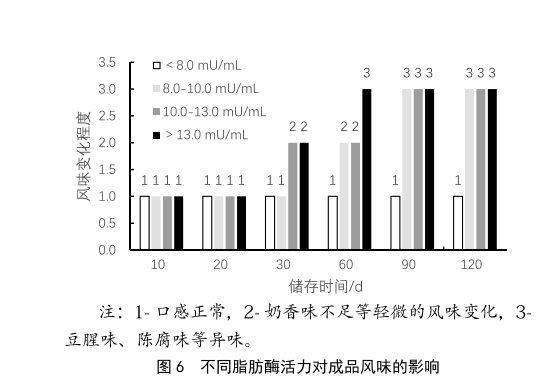
UHT产品由于其经过了超高温灭菌工艺,经过无菌灌装后,产品内部实现商业无菌状态,因此UHT产品可在常温下储存,且保质期较长,通常可达到6个月。但UHT产品在储存时经常会出现脂肪上浮、风味异常、口感变差等情况,这类情况会严重影响产品的货架期,并影响消费者的购买体验。原料奶中产生的耐热脂肪酶在经过超高温灭菌后,仍然不能被完全灭活,这部分脂肪酶会持续分解乳脂肪,产生脂肪酸,引起脂肪上浮或氧化味[5]。本次实验选取不同梯度的脂肪酶活力的原料奶,将其对应的UHT成品进行常温储存,模拟货架陈列。感官品评小组在10 d、20 d、30 d、60 d、90d和120 d后进行口感品评,客观的记录其风味,其感官结果见图6。
3 结论
牛乳中嗜冷菌代谢的耐热脂肪酶不会被超高温灭菌工艺完全灭活,残留的脂肪酶会持续分解成品乳中的脂肪,缩短产品货架期[6]。经实验表明,原料奶中脂肪酶活力越高,对应成品的风味变化越快,越容易产生脂肪氧化的豆腥味或陈腐味。原料奶中的脂肪酶活力随着储运时长的增加而不断提高,在72 h后降低,即使在轻微脱冷的情况下,脂肪酶活力的上升速度也会明显提高,所以在供应链系统中缩短运输、储存时间是控制脂肪酶活力的有效手段,对奶车的运输和原奶的储存温度的监控也至关重要。另外,供应链上游的奶源基地的卫生条件直接影响了牛乳中的微生物指标,奶源基地良好的卫生条件是保证原料奶中较低嗜冷菌数量和较低脂肪酶活力的必要条件。
参考文献
[1]张甦.哈尔滨地区生鲜牛乳中嗜冷菌数量与蛋白酶酶活力关系的测定[J].中国乳品工业,2012,40(7):41-42.
[2]史玉东,康小红.浅析UHT乳中耐热酶与产品质量问题的关系[J].农产品加工(学刊),2008(2):79-81.
[3]王欢,何腊平,周换景,等.脂肪酶活力测定方法及其在筛选产脂肪酶微生物中的应用[J].生物技术通报,2013(1):203-208.
[4]张爱霞.耐热酶与乳成分对UHT乳稳定性影响的研究[D].保定:河北农业大学,2003.
[5]郭德军,孙爱民,纪振杰.原料乳中嗜冷菌及其主要热稳定性酶类的研究进展[J].中国乳品工业,2006(8):46-49.
[6]胥伯强.嗜冷菌对长货架期乳制品的影响[J].中国食品工业,2003(3):50.






热点推荐
-

“喂饱”体内的益生菌,肠道更健康
-

全十红红稗饼干|中秋团圆,回家必备
-

和汪氏蜂蜜共同来普及蜂蜜结晶现象
-

拿坡海开启西餐加盟新潮流, 大众化家庭小西餐成未来新趋势
-

摩购空间“霸王餐”来袭 打破传统营销套路带动商家业绩翻倍
-
牛乳中脂肪酶活力检测方法与影响因素研究
-

果蔬中农药残留快速检测的质量控制探讨
-

气相色谱-质谱联用法测定绿茶中4种菊酯类杀虫剂农药残留量
-

几种常见食品中亚硝酸盐含量的测定
-
微波消解-电感耦合等离子体质谱法测定大米中的砷、铅、镉、铬
-
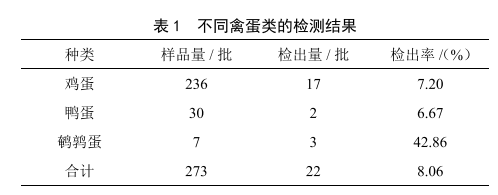
2019—2020年北海市禽蛋中6种兽药残留检测结果分析
-
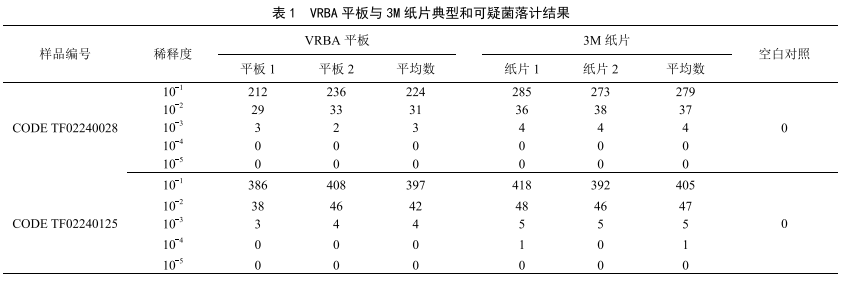
食品中大肠菌群检测结果与分析
-
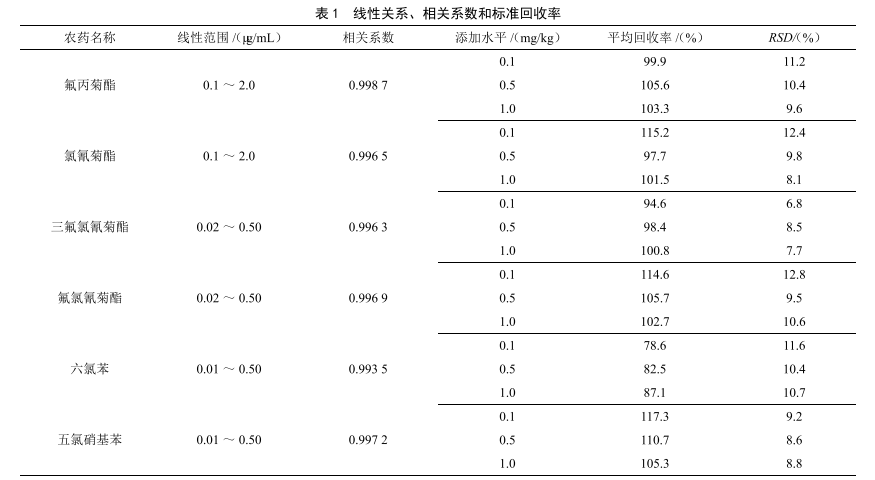
气相色谱法测定大米中多种农药残留量
-
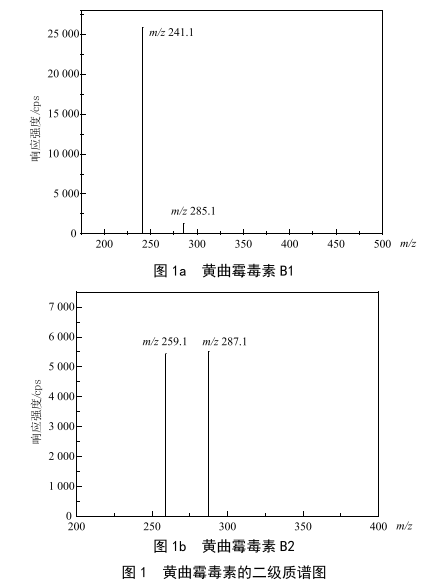
分散液液微萃取-超高效液相色谱-串联质谱法测定茶饮料中黄曲霉毒
-

辣椒及其制品中辣椒素含量检测及辣度分级
-
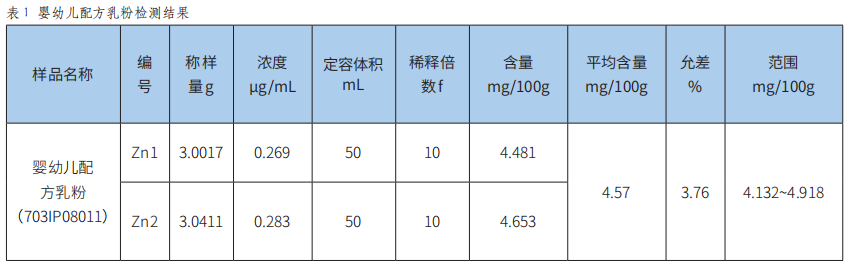
火焰原子吸收光谱法测定食品中锌的方法验证
-
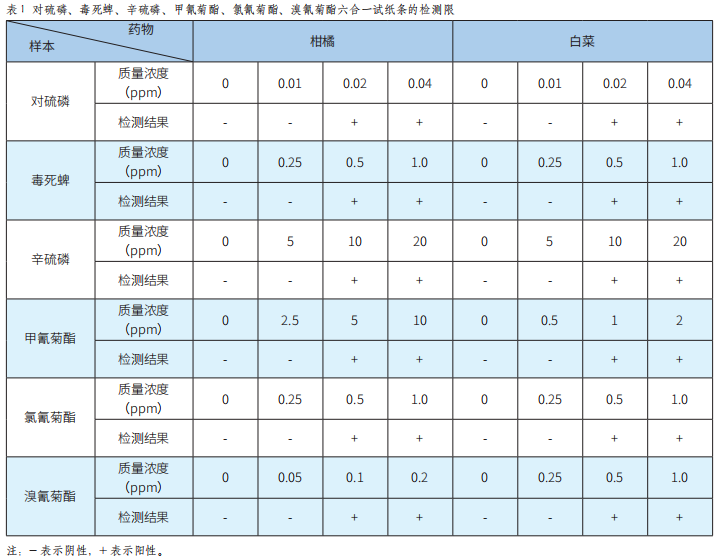
对硫磷等6种农药残留荧光免疫检测芯片的研发及应用
-

山东省潍坊市市售蔬菜中农药残留现况及慢性膳食暴露风险评估
-

食品检验检测机构质量控制的探讨
-
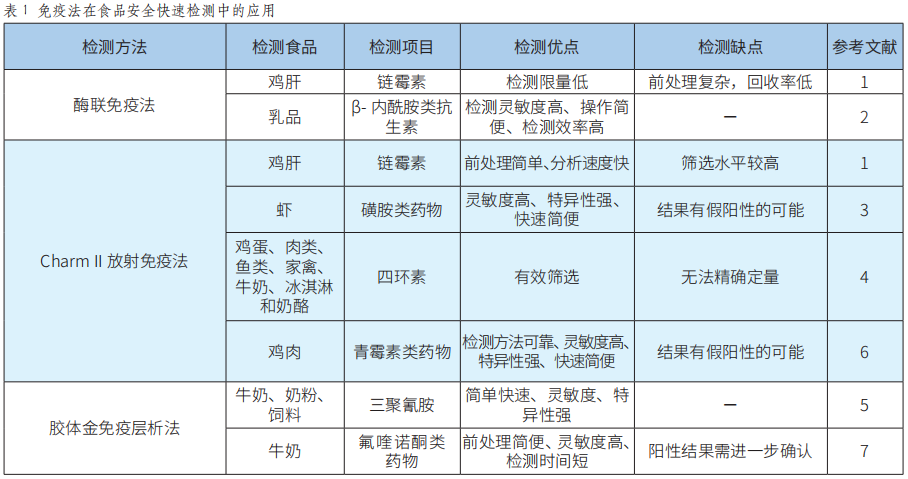
快速检测技术在食品安全中的应用研究进展
-
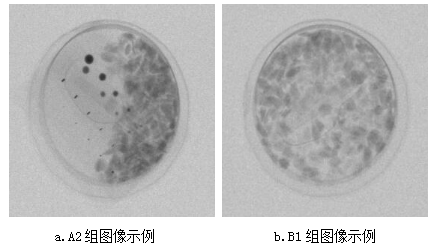
基于X射线图像的坚果盒异物检测
-
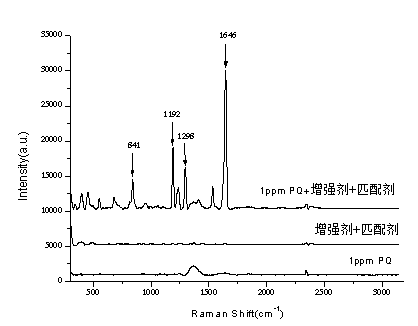
表面增强拉曼光谱法快速检测调味品中的百草枯
-
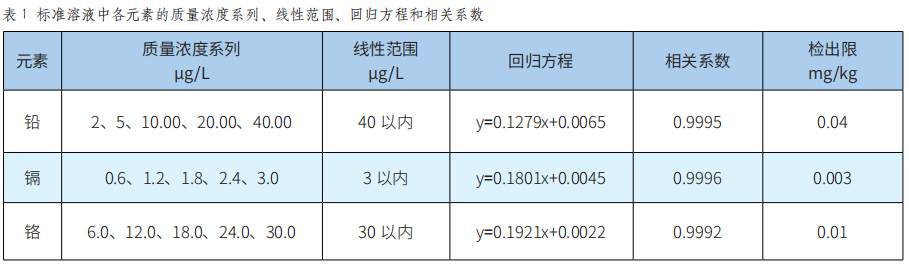
潞党参重金属污染物含量的评估
-
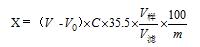
电位滴定法测定特殊医学用途食品中氯化物的不确定度评定
-
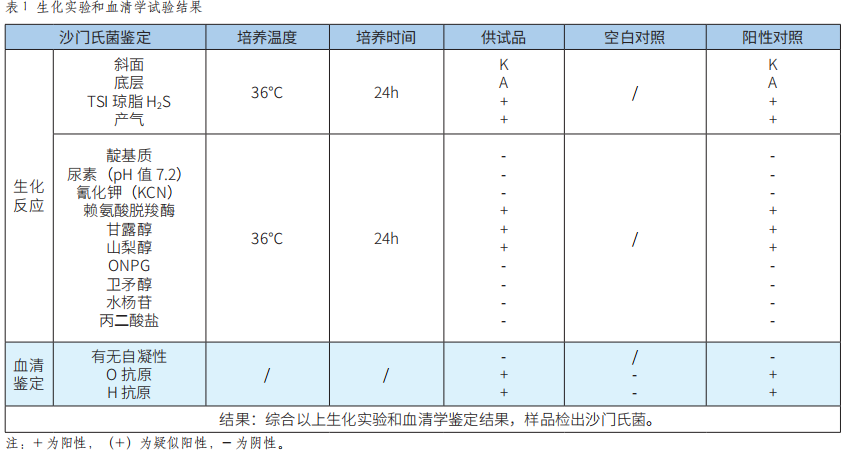
API 20E鉴定系统等两种检测方法应用于食品中沙门氏菌的检测结果
-

2020年上海市青浦区肉制品中单增李斯特菌分子分型及耐药性分析
-
一种基于ATP荧光反应的洁净度检测系统的开发与验证
-
固相萃取-高效液相色谱-二极管阵列检测法测定饮料中9种人工合成着
-

一种检测盐霉素胶体金试纸条的研制及其应用
-

高效液相色谱法测定绿豆糕中5种常见的食品添加剂